更新日:2022/2/4
免疫学問題集(1999年)
★作成時点での情報・記事であり,最新の情報ではありません。
1. 武装化Th1細胞によるマクロファージの活性化についてまとめよ。(CD4,ペプチド,MHC class II,IFNγ,CD40,Fasリカンド,TNFα,TNFβ,IL-3,GM-CSF,iNOS,NO,Oxygen
radicals,Pneumocystis carinii,intracellular pathogensなどのkeywordsを用いて説明せよ。以下同様)
- (解答例1)
- 細菌を貪食したマクロファージはMHC class IIに抗原を提示し,CD4 T細胞はTh1に分化・増殖する。このTh1はマクロファージを活性化する。マクロファージ活性化により殺される代表例はPneumocystis
cariniiである。エイズ患者ではCD4 T細胞が傷害されるため,Pneumocystis carinii感染が致命的となる。マクロファージ活性化には次の2つのシグナルが必要である。
1)IFNγ
2)CD40リガンドとTNFα,TNFβ
活性化されたマクロファージ内では,活性酸素・NOが産生され殺菌能が亢進する。NOはNO合成酵素(iNOS)により産生されるが,IFNγ,TNFαの相乗作用で産生が高まる。また,Th1は他にIL-3,GM-CSFを分泌し,骨髄でマクロファージ新生を促したり,細菌に慢性感染され活性化されなくなったマクロファージに対してFasリガンドを発現し,Fasを発現する細胞を破壊する。生体内でのマクロファージ活性化は,活性酸素やNOによる局所組織の破壊も伴うために,Th1により厳密にコントロールされている。
- (解答例2)
- ナイーブCD4 T細胞から分化したTh1細胞は,MHC class II分子に会合したペプチド抗原を認識する。Th1細胞は感染マクロファージに出会うと,IFNγ,GM-CSF,TNFα,CD40リガンド,Fasリガンドのようなマクロファージ活性化因子やIL-2,IL-3,TNFβなどを分泌する。
●IFNγとGM-CSF
マクロファージを活性化し,細胞内細菌を破壊する。
●FasリガンドやTNFβ
慢性感染に冒されたマクロファージを破壊し,排出された細菌を新たなマクロファージに破壊させる。
●IL-2
ナイーブT細胞をエフェクタ細胞へと分化増殖させる。
●IL-3とGM-CSF
骨髄でのマクロファージの分化を誘導する。
●TNFα
マクロファージを感染部位へ移行させる。
これらの因子によって活性化されたマクロファージは,CD40,TNFレセプターの発現が上昇し,TNFαも分泌する。これらの分子は,Th1細胞の分泌するIFNγと相乗的に作用し,NOや活性酸素(O・2)の産生を誘導する(NOはiNos(=
inducible NO syntase)という酵素が作る)。
2. 武装化CTLの働きを説明せよ。(CD8,MHC class I,ペプチド,パーフォリン,グランザイム,Fasリガンド,IFNγ,TNFα,TNFβ)
武装化CTLは宿主細胞内に寄生する病原体に効果を持つ。ナイーブCD8 T細胞は,病原体に由来するペプチド抗原とMHC class I分子の複合体を認識し,細胞障害性T細胞(CTL)へと分化する。あらかじめ傷害顆粒内に貯蔵されている2種のサイトトキシン(パーフォリン・グランザイム)の分泌によりアポトーシスを誘導する。アポトーシスはFasリガンドを介しても行われる。IFNγ,TNFα,TNFβを産生する。IFNγはウイルス複製を阻害し,MHC
class I分子の発現を増強することでCTLに攻撃されやすくする。TNFα,TNFβはIFNγと相乗的に働く。CTLは特異抗原を発現するものを選択的に傷害するため,近くの非感染細胞は傷害されない。
3. 武装化Th2細胞の働きを説明せよ。(B細胞活性化,CD40リカンド,IL-4,IL-5,IL-6ほか)
武装化Th2細胞は胸腺依存性の液性免疫応答に働く。B細胞に結合した抗原は細胞内に取り込まれ,細胞上にペプチド・MHC class II複合体として提示される。同一の抗原または病原体に対して感作された武装化Th2細胞はこれを特異的に認識し,CD40リガンドを表面に発現し,IL-4,5,6を放出する。CD40リガンドとIL-4は協調的にB細胞をクローン増殖させる。次にIL-5,6はB細胞を抗体分泌性の形質細胞へと分化させる。このようにしてB細胞は活性化される。
4. ナイーブT細胞からTh1,Th2への分化決定のしくみ。
ナイーブCD4 T細胞の分化は,病原体が誘導するサイトカインの種類によって決定される。細胞内寄生細菌やウイルスはマクロファージやNK細胞を活性化してIL-12やIFNγの産生を誘導する。IL-12やIFNγの存在化に活性化されたナイーブCD4
T細胞はTh1細胞に分化する。ある種の寄生虫はNK1.1+ CD4 T細胞に働いてIL-4を産生させる。IL-4の存在化に活性化されたナイーブCD4 T細胞はTh2細胞に分化する。すでにTh1またはTh2が存在するとき,これらのCD4
T細胞はお互いを抑制するサイトカインを産生する。活性化Th2細胞はTNFβとIL-10を産生し,Th1細胞の活性化と増殖を阻害する。活性化Th1細胞はIFNγを産生して,Th2細胞の増殖を阻害する。
5. 近交系マウスを用いた皮膚移植片拒絶実験により移植の成否を決定する遺伝子群の発見がもたらされた。これらの実験とその解釈を説明せよ。
近交系マウス:それぞれの個体の遺伝子構成が等しいとされているマウスたち
A系という同じ遺伝子配列を持ったマウスたちと,B系という同じ遺伝子配列を持ったマウスたちを使って実験する(A≠B)。
- 同じ系の中のマウス間で皮膚移植を行った場合は拒絶されないが,A-B間の移植では拒絶反応が起こる。
- A系マウス(AA)とB系マウス(BB)を交配した子孫F1(AB)の間において,(1)親からF1への移植は成功するが,(2)F1から親では拒絶反応が起こった。
- A系マウス(AA)と子孫F1(AB)を戻し交配した子孫F2(AA:AB=1:1)の半分はB系マウス(BB)の移植片を拒絶した。
この結果より次のことが分かる。
●2.(1)F1の免疫系は両親のどちらの抗原に対しても寛容である。
●2.(2)移植拒絶に関わる抗原のすべてがF1に発現している。
●単一の遺伝子座が急性移植片拒絶を支配している。
この遺伝子座は後に関連遺伝子の集団であることがわかり,主要組織適合遺伝子複合体(major histocompatibility,MHC)と名付けられた。MHCの発見により抗原性,細胞免疫,移植などの研究が進んだ。
6. 細菌感染に対する免疫応答をStaphylococcus aureusおよびMycobacterium tuberculosisを例にとり具体的に説明せよ。
- 1)Staphylococcus aureusに対する免疫応答
- 黄色ブドウ球菌は体内に侵入すると,マクロファージや好中球の食作用により取り込まれ,これらの細胞内で分解される。しかし黄色ブドウ球菌などのグラム陽性菌は莢膜を持っているため,なかなか取り込まれない。抗体や補体が細菌表面に結合すると,食細胞表面のFcレセプターに結合できるため,食作用が促進される(オプソニン作用)。貪食された細菌は食胞にリソソームが融合することにより分解され,ペプチド断片はMHC
class IIと複合体を作り,細胞表面に提示される。黄色ブドウ球菌は細胞外寄生細菌であるので,マクロファージはNK1.1細胞に働き,IL-4を産生させる。IL-4はナイーブT細胞CD4
T細胞をTh2細胞に分化させる。Th2細胞はB細胞を活性化し,体液性免疫を誘導する。
- 2)Mycobacterium tuberculosisに対する免疫応答
- 結核菌は体内に侵入すると,いったんはマクロファージによって貪食されるが,結核菌は食胞とリソソームの融合を阻害するためマクロファージ内で増殖する。マクロファージが細胞内の結核菌に由来するペプチドをMHC
class IIとともに細胞表面に発現すると,それを認識したナイーブCD4 T細胞はTh1細胞に分化する。Th1細胞はIFNγやCD40リガンドなどを産生し,マクロファージを活性化し,細胞内結核菌を破壊する。
7. ウイルス感染に対する免疫応答を説明せよ。(リンパ節,抗原提示細胞,CD8+ T細胞,CD4+ T細胞,B細胞,IgM抗体,IgG抗体,NK細胞,MHC
class I,MHC class II,IFNα/β,γの用語を用いて説明する)
ウイルスが体内に侵入すると,まずマクロファージによる非特異的な貪食が行われる。次に末梢で抗原を取り込んだマクロファージ,樹状細胞などの抗原提示細胞はリンパ節で捕捉され,ナイーブT細胞に抗原を提示し,エフェクタT細胞に分化させる。
細胞外のウイルスには,MHC class II・ペプチド複合体を認識したCD4+ T細胞はTh2となり,B細胞を活性化し,IgM抗体,IgG抗体を分泌させる。(液性免疫)
細胞に感染したウイルスには,(1)NK細胞・補体などによる非特異的な感染細胞の傷害(2)感染細胞のMHC class I・ペプチド複合体を認識したCD8+
T細胞はキラーT細胞となり,キラーT細胞による感染細胞の傷害が行われる。(細胞性免疫)
さらにインターフェロンによる増殖抑制作用も働く。
- ●IFNα/β
- 1)ウイルス複製の阻害
2)MHC class I分子の発現増強
3)NK細胞の増強・活性化
- ●IFNγ
- 1)〜3)
4)マクロファージ活性化
5)Th2を抑制し,Th1主体に誘導
8. 骨髄で生まれたBリンパ球が,抗体産生細胞になるまでの出来事を説明せよ。(骨髄,骨髄ストローマ細胞,抗原非依存性に免疫グロブリン遺伝子再編成(VDJ),未熟B細胞,自己抗原,クローン消失,抗原特異性,MHC,T-B相互作用,補助受容体,サイトカイン,胚中心,体細胞高頻度突然変異,アイソタイプスイッチなどを用いて説明)
骨髄の幹細胞から生まれたB細胞前駆体は,骨髄ストロマ細胞に依存しながら,抗原レセプターの多様性を得るため抗原非依存性に免疫グロブリン遺伝子再編成を行い,抗原特異性のあるレセプター(IgM)を表面に作る。
表面にIgMを発現した未熟B細胞のうち,自己抗原に反応するクローンは消失する。自己抗原に反応しないものはIgMとIgDを発現し,成熟B細胞となり末梢へ出現する。
抗原を取り込んだB細胞はリンパ組織のT細胞領域に捕捉され,MHC class II・ペプチド複合体として抗原を抗原特異的ヘルパーT細胞に提示し活性化する。B細胞は活性化Th2細胞により活性化されて増殖する。このときTh2細胞は表面にCD40リガンドを発現し,B細胞を増殖させるIL-4,IL-5,IL-6のサイトカインを分泌している。これらのサイトカインによりアイソタイプスイッチの一部が始まる。
活性化B細胞は近くの濾胞へ移動し,胚中心を作る。そこで早い増殖と体細胞高頻度突然変異を起こし,濾胞樹状細胞(FDC)上の抗原と高い親和性を持つものが選択され,低親和性のものはアポトーシスを起こす。高親和性B細胞はさらにT細胞と相互作用し,形質細胞または記憶B細胞へと成熟する。
9. MHC class I分子,MHC class II分子による抗原提示の仕組みを,発現する細胞,MHCの構成分子,どのようにペプチドが生成されるか,どのT細胞に提示するのかなどについて簡単に説明せよ。
MHC class I分子は,全ての有核細胞に発現している(赤血球には発現しない)。造血系細胞に多く発現している。膜を貫通するα鎖と貫通しないβ2ミクログロブリンからなるヘテロ2量体である。α鎖は3つのドメインからなる。
MHC class I分子によりCD8 T細胞に提示されるペプチドは,細胞質にある抗原蛋白である。これはlarge multifunctional protease(LMP)複合体により分解され,TAP分子を介して小胞体の内宮に輸送されたものに由来する。
MHC class II分子は通常,造血系細胞のサブセットと胸腺実質細胞にのみ発現している。2つの膜貫通糖蛋白鎖のαとβからなり,各鎖はそれぞれ2つのドメインからなる。
MHC class II分子によりCD8 T細胞に提示されるペプチドは,抗原提示細胞が細胞外より取り込み,エンドソームの中で蛋白分解酵素により分解されてできたものである。
10. T細胞受容体,補助受容体(CD28,CTLA4)からのシグナル伝達をIL-2遺伝子発現制御を例に説明せよ。またアナジーとは何か,どんな場合に重要となるのか。T細胞活性化を阻害するCsAやFK506の標的分子は何か,どのような作用をする分子か。
T細胞の活性化には,2つのシグナルが必要である。T細胞レセプター(TCR)と補助レセプターCD4が抗原提示細胞(APC)上のペプチド抗原・MHC class
II複合体と結合すると第1のシグナルが伝達される。このときAPC上のB7分子とT細胞のCD28が結合すると第2シグナルが伝達され,T細胞が活性化し,IL-2遺伝子発現とクローン増殖が起こる。
一方,T細胞が活性化するとCTLA-4分子が発現する。CTLA-4分子はAPCのB7分子と結合すると,活性化T細胞を抑制するシグナルを送り,IL-2の産生とT細胞の増殖が抑制される。
T細胞の活性化には,2つのシグナルが必要であるが,CD28を介した第2シグナル(補助刺激シグナル)が伝達されないとき,T細胞は活性化せず不応状態に陥る。この状態をアナジーといい,免疫系が自己抗原に反応するのを防ぐために重要な役割を果たしている。
CsA(シクロスポリンA)やFK506といった免疫抑制剤はT細胞のシグナルに影響を与えている。それぞれ,シクロフィリン,FK結合蛋白に結合する。CsAやFK506と結合したこれらの標的分子は,T細胞のcalcineurinと複合体を作り,転写因子の活性化を阻害する。
11. 2種類のヘルパーT細胞(Th1,Th2)分化決定のしくみの概略を種々作製された遺伝子破壊マウスの研究結果との関連も含めて説明せよ。
IFNγ遺伝子破壊マウスは感染のない状態では健康であるが,細胞内寄生細菌である結核菌などを感染させると致死的になる。これはマクロファージの活性化が起こらないことによる抗菌活性(NO産生)の低下と推測されている。またNK活性の低下も見られるなど細胞性免疫の低下が確認されている。この結果からIFNγが細胞内寄生細菌の排除に必要であることが分かった。また,IFNγ遺伝子破壊マウスは抗原刺激に対してTh1の生成が悪く,Th2生成が亢進している。その結果IL-4やIgEの産生が亢進する。
12. プロフェッショナル抗原提示細胞の性状と役割について説明せよ。
プロフェッショナル抗原提示細胞とは,樹状細胞・マクロファージ・B細胞のことをいう。これらの抗原提示細胞(APC)は,末梢で抗原を取り込んだ後,リンパ組織へ遊走し,そこで循環しているナイーブT細胞に抗原を提示する。
T細胞レセプター(TCR)がAPC上のペプチド抗原・MHC分子複合体と結合すると,補助レセプターのCD4を介するシグナルがT細胞に伝えられる(第1シグナル)。このとき,B7分子とCD28の結合を介する第2シグナル(補助刺激シグナル)が伝達されるとT細胞は活性化される。
活性化されたナイーブT細胞は,IL-2を分泌し,これに反応し増殖する。そして特異抗原による刺激のみでエフェクタ効果を発揮できる活性化エフェクタT細胞へ分化する。
13. サイトカイン受容体からのシグナルおよびシグナルにより制御される遺伝子群によりサイトカイン作用を説明せよ。(IL-2,IL-3,IL-6,IFN-α/β,IFN-γのいずれか)
- サイトカインの作用
- サイトカインは細胞から産生され,他の細胞の特異的レセプターに結合して細胞の挙動に影響するタンパク質で,細胞間にシグナルを伝えるシグナル伝達物質の1つである。
- シグナルの伝達
- (1)サイトカインが細胞上のレセプターに結合すると,レセプターの細胞内部分と結合しているプロテインキナーゼ群が,相互にリン酸化して活性化する。これらのリン酸化酵素はJanusキナーゼファミリーに属する。活性化したJanusキナーゼは,STATと呼ばれる遺伝子発現制御蛋白群をリン酸化し活性化する。
(2) リン酸化したSTATは,他のSTATや細胞質内蛋白と複合体を形成し,これが核内に移行してDNA上の特定の塩基配列に結合し,それぞれの遺伝子を活性化させる。
- IFNα/β
- ●作用1:ウイルスmRNAの分解
IFN + receptor →DNA→ 2-5A(オリゴアデニル酸)合成酵素→ウイルス2本鎖RNA+ATP→2-5Aの合成→RNaseの活性化→ウイルスmRNAの分解
●作用2:ウイルス蛋白合成の開始を阻害
IFN + receptor →DNA→プロテインキナーゼ→ウイルス2本鎖RNA+ATP→ウイルス蛋白合成開始因子のリン酸化→ウイルス蛋白合成開始を阻害
※IFNが細胞のレセプター(CD118)に結合すると,レセプターの細胞内部分と結合しているJAK1やTyk2などのJanusキナーゼが相互にリン酸化し活性化する。活性化したJanusキナーゼは,STAT1-4(遺伝子発現制御蛋白群)をリン酸化し活性化する。リン酸化したSTATは,他のSTATや細胞質内蛋白と複合体を作りこれが核内に移行してISREに結合しているp48と3量体を形成して遺伝子を活性化させ,それぞれの機能を発現させる。
14. 抗H-Y抗原に対するT細胞受容体遺伝子を導入したトランスジェニックマウスの実験により,胸腺内ポジティブセレクション,ネガティブセレクションの存在が証明されたが,この実験系の概要と得られた結果の要約,結論を述べよ。
- *1トランスジェニックマウス
- 外来性遺伝子を組み込んで次世代でその遺伝子を発現するようにされたマウスで,導入遺伝子の機能解析に役立つ。
- *2ポジティブセレクション
- ペプチド抗原+MHCと結合力の弱いTCRを持つT細胞が選択され,分化増殖し,末梢へ移行する。
- *3ネガティブセレクション
- ペプチド抗原+MHCと結合力の強いTCRを持つT細胞はアポトーシスを起こす。
まず,マウス♂に特異的なH-Y抗原を認識し,bというMHCに拘束されるTCR遺伝子を導入したトランスジェニックマウスを作成する。
(1)b-MHCの♀では導入したTCRを発現した成熟T細胞が胸腺や末梢リンパ節に大量に見つかった。
(2) d-MHCの♀では導入したTCRのMHCはb拘束性なので,MHCに反応できない。末梢に導入したTCR遺伝子を発現した成熟T細胞はほとんど存在しなかった。
(3) b-MHCの♂では導入したTCRのリガンドであるH-Y抗原とともに反応できるb?MHCが発現していて,胸腺の細胞数が激減し,導入したTCRを発現した成熟T細胞は胸腺・末梢リンパ節ともに激減していた。
(1),(2)より♀では胸腺内でポジティブセレクションが,(3)より雄ではネガティブセレクションが起きていることが明らかになった。
(参考)ドナー,レシピエントのMHCが一致しても他の遺伝子座が異なっているとき拒絶反応が起きる(当然だけど)。MHCの一致した移植の拒絶に関するMHC多型はマイナーH抗原(副組織適合抗原:minor
histocompatibility antigen)と呼ばれる。
マイナーH抗原に対する反応を誘導する蛋白群はY染色体にコードされており,H-Yと呼ばれる。このため女性では抗男性マイナーH反応が起こる。
マイナーH抗原は自己ペプチドとして移植片上の全ての細胞でMHC class I分子により提示されるので,自己免疫により移植片全体が破壊される。このために移植の際には免疫抑制剤を投与する。
15. 細胞障害性T細胞の活性化と機能について知るところを記せ。
ナイーブCD8 T細胞はあらかじめキラーT細胞に分化することになっている。ナイーブT細胞の活性化には(1)特異抗原シグナルと(2)補助刺激シグナルの2つが必要である。(1)特異抗原シグナルは,抗原提示細胞のMHC
class I+抗原ペプチドとナイーブT細胞のTCR+補助レセプターCD8が結合することで伝えられる。(2)補助刺激シグナルは,抗原提示細胞のB7分子とナイーブT細胞のCD28分子が結合することでT細胞に送られる。
これらの2つのシグナルによってはじめてナイーブT細胞は活性化されて,IL-2を分泌し,自身のIL-2に反応して増殖・分化するが,(1)のシグナルのみだと抗原で感作されても活性化されないアナジー状態となる。また,いったん武装化エフェクタT細胞になると(1)のシグナルのみでエフェクタ効果を発揮し,(2)のシグナルは必要としない。
活性化したキラーT細胞は,標的細胞上のMHC class I分子+抗原ペプチドを認識すると,標的細胞にアポトーシスを起こさせるが,その方法はあらかじめ産生された2種類のサイトトキシン(パーフォリン・グランザイム)を分泌することによる。グランザイムは標的細胞にアポトーシスを起こさせる。パーフォリンは標的細胞の細胞質膜に小孔を形成し,グランザイムの侵入を助ける。また,膜結合性分子のFasリガンドを発現し,標的細胞のFasに結合させ,標的細胞にアポトーシスを起こさせる。さらにIFNγを産生し,ウイルス増殖の阻害,MHC
class I分子の発現増強およびマクロファージの活性化を行う。
16. アポトーシスはどんな場合に重要な役割を果たしているのか。それぞれの場合を示し意義を論じよ。
- B細胞
- 1)骨髄:自己反応性B細胞は消去され,自己寛容性を獲得する。
2)末梢:(1)自己抗原に反応し,アネルギー状態になったB細胞はCD4 T細胞からFasを介し消去される。(2)胚中心で体細胞高頻度突然変異によって,a)自己抗原に反応するものやb)抗原親和性が低いものは消去される。
- T細胞
- 胸腺における分化・成熟過程で1)自己MHCを認識しないもの(正の選択)や2)自己抗原に反応するもの(負の選択)は消去される。
- エフェクタT/B細胞
- 記憶細胞にならなかったものはアポトーシスを起こす。→免疫反応の停止
- CTL
- 1)パーフォリン・グランザイムを分泌して標的細胞を殺す。
2)Fasを発現した細胞を殺す→感染細胞内病原体のDNAを破壊し,近隣細胞への感染を阻止する。
17. 自然免疫と獲得免疫について簡潔に説明せよ。自然免疫に関わる細胞,液性因子にはどんなものがあるのか。
- 1)自然免疫
- 生まれつき備わっている免疫で,感染に対する第一線の防御機構で,抗原非特異的かつ抗原の侵入に対して速やかに機能する。免疫学的記憶は生じない。
関わる細胞:マクロファージ,好中球,NK細胞
液性因子:補体,ミエロペルオキシダーゼ,ムラミダーゼ,MBP(major basic protein),ヒスタミン
- 2)獲得免疫
- 抗原特異的で免疫学的記憶を生じるため,再感染に対しては速やかかつ強力な免疫反応が起こり,発症は未然に防がれる。リンパ球の働きによる。
18. アレルギーの4つの型を説明せよ。
- 1)I型
- ある抗原に対するIgE抗体が肥満細胞や好塩基球のFcレセプターに結合し,抗体に対する抗原が結合すると,細胞内顆粒内のヒスタミン・セロトニンが放出される。これらの物質はアナフィラキシー症状(鼻炎・喘息)を起こす。
- 2)II型
- 薬剤などが細胞表面に結合すると,IgGやIgMの抗体が結合し,補体の活性化・マクロファージなどによる破壊が起こる。溶血性貧血,糸球体腎炎,心内膜炎などを起こす。
- 3)III型
- 抗原と抗体が結合して免疫複合体を作り,それが組織に沈着すると,補体の活性化が起こる。肥満細胞の脱顆粒により血管透過性の亢進,白血球の遊走,凝固系の活性化などが起こり,周囲の組織が傷害される。
- 4)IV型
- エフェクタT細胞は,エフェクタ分子を放出したり(Th1),抗原に再刺激され細胞を傷害する(CTL)。この反応は,エフェクタ分子を産生するため時間がかかり,遅延型過敏反応と呼ばれる。
19. T細胞やB細胞の活性化を制御する分子群として活性化阻止分子群が知られている。これら分子群の役割と作用機序を説明せよ。
20. APC,B細胞,ヘルパーT細胞相互作用について説明せよ。
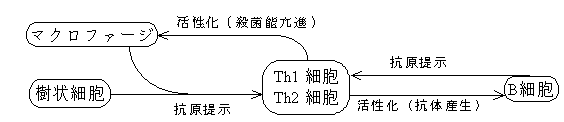
APC(マクロファージ,樹状細胞,B細胞)はヘルパーT細胞にMHC class IIを介して抗原提示を行い,ナイーブCD4 T細胞をTh1,Th2細胞に分化させる。
Th1細胞はマクロファージ活性化因子(IFNγとCD40リガンド)によりマクロファージを活性化し,殺菌能を亢進させる。
Th2細胞はB細胞の増殖因子(IL-4,IL-5,CD40リガンド)によりB細胞を増殖させ,形質細胞へ分化させる。
21. 胸腺におけるポジティブセレクション,ネガティブセレクションを説明せよ。
T細胞前駆細胞は胸腺で次の段階を通り成熟する。
| 被膜下 |
CD3-4-8- |
ダブルネガティブ胸腺細胞 |
| |
↓増殖 |
|
| 皮質 |
CD3+4+8+ |
ダブルポジティブ胸腺細胞 |
| |
↓正の選択・負の選択 |
|
| |
CD4+8-またはCD4-8+ |
シングルポジティブ胸腺細胞 |
| |
↓ |
|
| |
末梢への移動 |
|
正の選択は胸腺上皮細胞の働きによる。胸腺上皮細胞は表面に自己MHC分子を発現している。胸腺上皮細胞上のMHC分子がMHC class IのときはCD8分子,MHC
class IIのときはCD4分子を発現したT細胞のみが成熟する(自己MHC拘束性の獲得)。また胸腺上皮細胞表面の自己MHC分子を認識しないダブルポジティブ胸腺細胞(95%)はアポトーシスを起こして死んでしまう。
負の選択はマクロファージ,樹状細胞などの抗原提示細胞の働きによる。T細胞が抗原提示細胞上の自己MHCに結合している自己ペプチドに出会うと,これに特異的なTCRを持ったT細胞は消去される。このようにして自己抗原に反応しないT細胞のみが成熟するため自己寛容性を維持している。
22. NK細胞の働き(ウイルス感染,がん細胞に対する防御)を説明せよ。
NK細胞はリンパ系幹細胞から分化した細胞であるが,T/B細胞とは異なり,免疫刺激がなくても非自己,異常細胞を破壊できる。
ウイルス感染では感染初期にIFNα/β,TNFβ,IL-12などが産生され,NK細胞が活性化される。これらの作用によりウイルス増殖が抑制されるが,ウイルスを除去するためにはウイルス特異的CD8
T細胞が必要である。
NK細胞は次のようにして腫瘍細胞やウイルス感染細胞に対する細胞障害性を発揮する。NK細胞はNKR-PIというレセプターを持ち,自己細胞上の糖鎖を認識すると活性化して細胞障害性を発揮するが,第2のレセプター(マウスではLy49,ヒトではKIR:Killer
Inhibitary Receptor)がMHC class I分子に結合すると,NK細胞の細胞障害性は抑制される。もし宿主細胞がMHC class Iを発現しないときやMHCの形態が変化しているときは,抑制性シグナルが送られないので,活性化NK細胞による障害を受ける。
23. スーパー抗原とは何か。
通常T細胞の活性化には,MHCによる抗原ペプチドの提示が必要である(MHC拘束性)。しかし,ブドウ球菌のエンテロトキシンやTSST-1毒素のように,MHCの抗原提示を介さなくても強力にT細胞を活性化させる抗原が存在する。このような抗原ペプチドをスーパー抗原という。
スーパー抗原は通常の抗原提示機序に従わず,TCRのβサブユニットのVβ領域に直接結合することによりT細胞を活性化する。スーパー抗原は生体内で特殊な作用を受けることなくそのような効果を現すことができ,スーパー抗原ペプチドを断片化すると生理活性を失う。
24. HLAの多様性と抗原への応答のレパートリーとの関連を説明せよ。
近交系マウスの皮膚移植拒絶実験により,移植の成否を決定する遺伝子群としてMHCが発見されたが,ヒトにおいてヒトMHCは最初白血球細胞と反応する抗体を用いて研究されたため,ヒト白血球抗原human
leukocyte antigen:HLA系と呼ばれた。
ヒトは数多くの抗原に対応するために,MHC対立遺伝子の多様性を生み出し,MHCの多型性を獲得することが必要であった。なぜならMHCのペプチド収容溝における特定のアミノ酸側鎖を変異させることでより多くのペプチド抗原と結合でき,T細胞に提示できるようになるからである。
ヒトMHCをコードする遺伝子座は複数存在し,中には100以上もの対立遺伝子を持つものがあり,またこれらは共優性であるために発現するMHC分子の種類は2倍になる。この機構によりヒトMHC遺伝子に関して非常に多くの変異を有することになる。これらの変異のほとんどはペプチド収容溝の形の違いとなって現れる。この多型性が抗原のレパートリーの多さに対応している。
25. サイトカイン受容体とT細胞抗原受容体について細胞膜受容体から核内遺伝子発現制御に至る細胞内シグナル伝達経路をそこに関わる重要な分子群の活性化経路を示しながら説明せよ。
- 1)サイトカイン受容体(Immunobiology 4th ed. Fig5.21参照)
- サイトカイン受容体は少なくとも2本の鎖からなっており,細胞質内ドメインにはJAKs(Janus kinases)が結合している。サイトカインが受容体と結合すると,受容体は2量体となり,JAKsも一緒になり互いに活性化する。その後受容体末端のチロシンがリン酸化される。次に転写因子STATsがリン酸化した受容体に結合すると,活性化されたJAKsによってリン酸化される。リン酸化されたSTATsは2量体を形成し,速やかに核に移行する。そこで,STATsは適応免疫に重要な遺伝子の翻訳を活性化させる。
- 2)T細胞受容体(Immunobiology 4th ed. Fig5.12参照)
- T細胞レセプターにMHC・ペプチド複合体が結合するとFynのような受容体に連結しているキナーゼが活性化し,CD3のγ,d,eのITAMをz鎖と同様にリン酸化する。チロシンキナーゼZAP-70はリン酸化されたz鎖のITAMに結合する。補助レセプターが抗原提示細胞のMHC分子に結合すると,LckはZAP-70をリン酸化し活性化する。
26. 抗体の作用(中和,オプソニン化,補体の活性化)を説明せよ。
- 1)中和
- 抗体は毒素,細菌,ウイルスに結合することで,宿主細胞への傷害,感染を阻止する。
- 2)オプソニン化
- 抗体は病原体と複合体を形成すると,マクロファージなどのFc部位に結合できるようになり,食作用を促進する。
- 3)補体の活性化
- 抗体は補体を活性化し,それによって抗体のオプソニン化を促進し,細菌膜に穴をあけて破壊する。
27. IL-1,TNFα,IL-6の作用をまとめよ。
IL-1,TNFα,IL-6は非特異的な刺激により食細胞から分泌され,全身性の体温上昇作用と急性期反応の誘導作用を持つ。これらサイトカインは内因性発熱物質と呼ばれ,宿主の感染防御に役立っている。ほとんどの微生物は体温より低めの温度でよく増殖するが,適応免疫反応は高めの温度でよく機能する。また高温では宿主細胞はTNFαの悪い作用から免れることができる。IL-1,TNFα,IL-6は急性期蛋白を肝細胞に作らせ,好中球を作る骨髄を活性化する。急性期蛋白はオプソニンとして働き,オプソニン化された病原体は骨髄からの好中球に排除される。IL-1,TNFα,IL-6は視床下部,筋細胞,脂肪細胞に働き,体温上昇を誘導する。体温上昇により細菌やウイルスの増殖が抑制されるとともに,抗原のプロセシングや特異的免疫反応が亢進される。
【参照文献】
●免疫生物学 第3版
●ImmunoBiology 4th edition
|
